
核酸-k8 casino
核酸是脱氧核糖核酸(dna)和核糖核酸(rna)的总称,是由许多核苷酸单体聚合成的生物大分子化合物,为生命的最基本物质之一。 核酸
核酸
核酸核酸是一类生物聚合物,是所有已知生命形式必不可少的组成物质,是所有生物分子中最重要的物质,广泛存在于所有动植物细胞、微生物体内。
核酸由核苷酸组成,而核苷酸单体由五碳糖、磷酸基和含氮碱基组成。如果五碳糖是核糖,则形成的聚合物是rna;如果五碳糖是脱氧核糖,则形成的聚合物是dna。
概况简介 编辑本段
高分子化合物的一类,是生命的最基本物质之一。由许多(至少几十)个核苷酸通过磷酸二酯键连接而成,存在于所有动植物细胞、微生物和病毒、噬菌体内,对生物的生长、遗传、变异等现象起着决定作用。根据所含成分,可分为核糖核酸(rna)和脱氧核糖核酸(dna)两类。核酸与遗传、肿瘤的发生、病毒的感染性、射线对机体的作用都有重要关系。
种类作用 编辑本段
核酸同蛋白质一样,也是生物大分子。核酸的相对分子质量很大,一般是几十万至几百万。核酸水解后得到许多核苷酸,实验证明,核苷酸是组成核酸的基本单位,即组成核酸分子的单体。一个核苷酸分子是由一分子含氮的碱基、一分子五碳糖和一分子磷酸组成的。根据五碳糖的不同可以将核苷酸分为脱氧核糖核苷酸和核糖核苷酸。
核酸大分子可分为两类:脱氧核糖核酸(dna)和核糖核酸(rna),在蛋白质的复制和合成中起着储存和传递遗传信息的作用。核酸不仅是基本的遗传物质,而且在蛋白质的生物合成上也占重要位置,因而在生长、遗传、变异等一系列重大生命现象中起决定性的作用。
核酸 | dna | rna |
名称 | 脱氧核糖核酸 | 核糖核酸 |
结构 | 规则的双螺旋结构 | 通常呈单链结构 |
基本单位 | 脱氧核糖核苷酸 | 核糖核苷酸 |
五碳糖 | 脱氧核糖 | 核糖 |
含氮碱基 | a(腺嘌呤) g(鸟嘌呤) c(胞嘧啶) t(胸腺嘧啶) | a(腺嘌呤) g(鸟嘌呤) c(胞嘧啶) u(尿嘧啶) |
分布 | 主要存在于细胞核,少量存在于线粒体和叶绿体 | 主要存在于细胞质 |
功能 | 携带遗传信息,在生物体的遗传、变异和蛋白质的生物合成中具有极其重要的作用 | 作为遗传物质:只在rna病毒中;不作为遗传物质:在dna控制蛋白质合成过程中起作用。mrna是蛋白质是合成的直接模板、trna能携带特定氨基酸、rrna是核糖体的组成成分;催化作用:酶的一种 |
(1)五碳糖——dna是脱氧核糖;rna是核糖。
(2)碱基——dna是a、t、c、g(不含u);rna是a、u、c、g(不含t)。
(3)dna通常是双螺旋结构;rna通常是单链,局部可形成双螺旋结构。
核酸类似物是与天然存在的rna和dna类似(结构相似)的化合物,用于医学和分子生物学研究。核酸类似物在组成核酸的核苷酸分子以及组成核苷酸的碱基、五碳糖和磷酸基团的分子间发生了改变。通常,这些改变使得核酸类似物种的碱基配对和碱基堆积性质发生了改变。比如通用碱基可与所有四个经典碱基配对,又比如磷酸-糖骨架类似物(如pna)甚至可形成三重螺旋。核酸类似物也称为异种核苷酸,代表了异种生物学的主要支柱之一,即基于替代生物化学的新生自然形式的生命设计。
核酸类似物包括肽核酸(pna),吗啉代和锁核酸(lna)以及乙二醇核酸(gna)和苏糖核酸(tna)。因为分子主链发生了改变,它们与天然存在的dna或rna有明显的不同。
核酸在实践应用方面有极重要的作用,现已发现近2000种遗传性疾病都和dna结构有关。如人类镰刀形红血细胞贫血症是由于患者的血红蛋白分子中一个氨基酸的遗传密码发生了改变,白化病患者则是dna分子上缺乏产生促黑色素生成的酪氨酸酶的基因所致。肿瘤的发生、病毒的感染、射线对机体的作用等都与核酸有关。70年代以来兴起的遗传工程,使人们可用人工方法改组dna,从而有可能创造出新型的生物品种。如应用遗传工程方法已能使大肠杆菌产生胰岛素、干扰素等珍贵的生化药物。
核酸的作用
dna是储存、复制和传递遗传信息的主要物质基础。
rna在蛋白质合成过程中起着重要作用——其中转运核糖核酸,简称trna,起着携带和转移活化氨基酸的作用;信使核糖核酸,简称mrna,是合成蛋白质的模板;核糖体的核糖核酸,简称rrna,是细胞合成蛋白质的主要场所。
此外,现在已知许多其他种类的功能rna,如microrna等。
核酸类似物主要用于医学和分子生物学研究。
研究历史 编辑本段
1869年,f.miescher从脓细胞中提取到一种富含磷元素的酸性化合物,因存在于细胞核中而将它命名为“核质”(nuclein)。但核酸(nucleic acids)这一名词在miescher发现“核质”20年后才被正式启用,当时已能提取不含蛋白质的核酸制品。早期的研究仅将核酸看成是细胞中的一般化学成分,没有人注意到它在生物体内有什么功能这样的重要问题。[1]
在19世纪80年代早期,德国生物化学学家,1910年诺贝尔生理和医学奖获得者科塞尔进一步纯化获得核酸,发现了它的强酸性。他后来也确定了核碱基。
1889年,德国病理学家richard altmann创造了核酸这一术语,取代了nuclein。
1919年,一位美籍俄罗斯医生和化学家菲巴斯·利文首先发现了单核苷酸的三个主要成分(磷酸盐、戊糖和氮基)的顺序。
1938年,英国物理学家和生物学家威廉·阿斯特伯里和florence bell(后来改名为florence sawyer)发表了第一个dna的x射线衍射图谱。
核酸的实验研究构成了现代生物学和医学研究的重要组成部分,并为基因组和法医学以及生物技术和制药工业奠定了基础。
1944年,avery等为了寻找导致细菌转化的原因,他们发现从s型肺炎球菌中提取的dna与r型肺炎球菌混合后,能使某些r型菌转化为s型菌,且转化率与dna纯度呈正相关,若将dna预先用dna酶降解,转化就不发生。结论是:s型菌的dna将其遗传特性传给了r型菌,dna就是遗传物质。从此核酸是遗传物质的重要地位才被确立,人们把对遗传物质的注意力从蛋白质移到了核酸上。
核酸研究中划时代的工作是watson和crick于1953年创立的dna双螺旋结构模型。模型的提出建立在对dna下列三方面认识的基础上:
1.核酸化学研究中所获得的dna化学组成及结构单元的知识,特别是chargaff于1950~1953年发现的dna化学组成的新事实;dna中四种碱基的比例关系为a/t=g/c=1。
2.x线衍射技术对dna结晶的研究中所获得的一些原子结构的最新参数。
3.遗传学研究所积累的有关遗传信息的生物学属性的知识。综合这三方面的知识所创立的dna双螺旋结构模型,不仅阐明了dna分子的结构特征,而且提出了dna作为执行生物遗传功能的分子,从亲代到子代的dna复制 (replication)过程中,遗传信息的传递方式及高度保真性。其正确性于1958年被meselson和stahl的著名实验所证实。dna双螺旋结构模型的确立为遗传学进入分子水平奠定了基础,是现代分子生物学的里程碑。从此核酸研究受到了前所未有的重视。
dna分子具有规则的双螺旋结构。是由两条相互平行且反向右旋的脱氧核苷酸长链所构成,分子中央的碱基碱基互补配对原则以氢键相连。dna独特的双螺旋结构和碱基互补配对能力使dna的两条链“可分”,“可合”,半保留复制自如,“精确”复制的dna通过细胞分裂等方式传递下去,使子代(或体细胞)含有与亲代相似的遗传物质。但“精确”复制并不是绝对不存在差错,复制差错率非常低(约1~10亿分之一),然而却导致基因发生突变,出现新基因,产生可遗传的变异,有利于生物的进化。
瑞士生物学家:米舍尔friedrich miescher
美国生物学家:沃森watson,james dewey
英国生物物理学家:克里克crick,francis harry compton
英国医生:格里菲思griffith, f.
阿委瑞avery o. t.
赫尔希hershey, a.d.
蔡斯chase, m.
30多年来,核酸研究的进展日新月异,所积累的知识几年就要更新。其影响面之大,几乎涉及生命科学的各个领域,现代分子生物学的发展使人类对生命本质的认识进入了一个崭新的天地。
双螺旋结构创始人之一的crick于1958年提出的分子遗传中心法则(centraldogma),揭示了核酸与蛋白质间的内在关系,以及rna作为遗传信息传递者的生物学功能。并指出了信息在复制、传递及表达过程中的一般规律,即dna→rna→蛋白质。
遗传信息以核苷酸顺序的形式贮存在dna分子中,它们以功能单位在染色体上占据一定的位置构成基因(gene)。因此,搞清dna顺序无疑是非常重要的。1975年 sanger发明的dna测序(dnasequencing)加减法为实现这一企图起了关键性的作用。由此而发展起来的大片段dna顺序快速测定技术──maxam 和gilbert的化学降解法(1977年)和sanger的末端终止法(1977年),已是核酸结构与功能研究中不可缺少的分析手段。
中国学者洪国藩于1982年提出了非随机的有序dna测序新策略,对dna测序技术的发展作出了重要贡献。凭借先进的dna测序技术及其它基因分析手段,人类正在进行一项以探明自身基因组(genome)全部核苷酸顺序(单倍基因组 含3×10碱基对)为目标的宏伟计划──人类基因组图谱制作计划(human genome mapping project)。据称,此项计划的实现,将对全人类的健康产生无止境的影响。
watson-crick模型创立36年后的1989年,一项新技术──扫描隧道显微镜(scanning tummeling microscopy,stm)使人类首次能直接观测到近似自然环境中的单个dna分子的结构细节,观测数据的计算 机处理图像能在原子级水平上精确度量出dna分子的构型、旋转周期、大沟(major groove)及小沟(minor groove)。这一成果是对dna 双螺旋结构模型真实性的最直接而可信的证明。此项技术无疑会对人类最终完全解开遗传之谜提供有力的帮助。可喜的是,中国科学家在这项世界领先的研究中也占有一席之地。
大小组成 编辑本段
核酸分子通常很大。实际上,dna分子可能是已知的最大的单个生物分子。
但也有比较小的核酸分子。
核酸分子的大小范围从21个核苷酸(小干扰rna)到大染色体(人类染色体是一个含有2.47亿个碱基对的单个分子)不等。
核酸完全水解产生嘌呤和嘧啶等碱性物质、戊糖(核糖或脱氧核糖)和磷酸的混合物。核酸部分水解则产生核酸和核苷酸。每个核苷分子含一分子碱基和一分子戊糖,一分子核苷酸部分水解后除产生核苷外,还有一分子磷酸。
dna和rna含有的核糖同,dna含有脱氧核糖,而rna含有核糖。此外,dna和rna中含有的碱基也有差别:dna和rna都含有腺嘌呤,胞嘧啶和鸟嘌呤,但dna中不含有尿嘧啶,只有胸腺嘧啶
核酸中的糖和磷酸盐通过磷酸二酯键以交替链(糖-磷酸骨架)相互连接。磷酸基团所连接的碳是糖的3'-末端,与碳原子结合的碳是5'-末端,这就产生了核酸的方向性。核碱基通过n-糖苷键与糖连接。
在rna和dna中也发现了非标准核苷,它们通常来自dna分子内的标准核苷或初始rna转录物的修饰。转移rna(trna)分子含有特别多的修饰核苷。
天然存在的dna分子在大多数情况下是双链的,而rna分子是单链的。然而,有许多例外。一些病毒具有由双链rna构成的基因组,而其他病毒具有单链dna基因组,并且在某些情况下,可形成具有三个或四个链的核酸结构。
化学成分 编辑本段
核酸是生物体内的高分子化合物。它包括脱氧核糖核酸(deoxyribonucleicacid,dna)和核糖核酸(ribonucleicacid,rna)两大类。
单个核苷酸是由含氮有机碱(称碱基)、戊糖(即五碳糖)和磷酸三部分构成的。
碱基(base):构成核苷酸的碱基分为嘌呤(purine)和嘧啶>;(pyrimi-dine)二类。前者主要指腺嘌呤(adenine,a)和鸟嘌呤(guanine,g),dna和rna中均含有这二种碱基。后者主要指胞嘧啶(cytosine,c)胸腺嘧啶(thymine,t)和尿嘧啶(uracil,u),胞嘧啶存在于dna和rna中,胸腺嘧啶只存在于dna中,尿嘧啶则只存在于rna中。
嘌呤环上的n-9或嘧啶环上的n-1是构成核苷酸时与核糖(或脱氧核糖)形成糖苷键的位置。
此外,核酸分子中还发现数十种修饰碱基(themodifiedcomponent),又称稀有碱基,(unusualcomponent)。它是指上述五种碱基环上的某一位置被一些化学基团(如甲基化、甲硫基化等)修饰后的衍生物。一般这些碱基在核酸中的含量稀少,在各种类型核酸中的分布也不均一。如dna中的修饰碱基主要见于噬菌体dna,rna中以trna含修饰碱基最多。
戊糖(五碳糖):rna中的戊糖是d-核糖(即在2号位上连接的是一个羟基),dna中的戊糖是d-2-脱氧核糖(即在2号位上只连一个h)。d-核糖的c-2所连的羟基脱去氧就是d-2脱氧核糖。
戊糖c-1所连的羟基是与碱基形成糖苷键的基团,糖苷键的连接都是β-构型。
核苷(nucleoside):由d-核糖或d-2脱氧核糖与嘌呤或嘧啶通过糖苷键连接组成的化合物。核酸中的主要核苷有八种。
核苷酸(nucleotide):核苷酸与磷酸残基构成的化合物,即核苷的磷酸酯。核苷酸是核酸分子的结构单元。核酸分子中的磷酸酯键是在戊糖c-3’和c-5’所连的羟基上形成的,故构成核酸的核苷酸可视为3’-核苷酸或5’-核苷酸。dna分子中是含有a,g,c,t四种碱基的脱氧核苷酸;rna分子中则是含a,g,c,u四种碱基的核苷酸。
当然核酸分子中的核苷酸都以形式存在,但在细胞内有多种游离的核苷酸,其中包括一磷酸核苷、二磷核苷和三磷酸核苷。
3’,5’-磷酸二酯键:核酸是由众多核苷酸聚合而成的多聚核苷酸(polynucleotide),相邻二个核苷酸之间的连接键即:3’,5’-磷酸二酯键。这种连接可理解为核苷酸糖基上的3'位羟基与相邻5'核苷酸的磷酸残基之间,以及核苷酸糖基上的5'位羟基与相邻3'核苷酸的磷酸残基之间形成的两个酯键。多个核苷酸残基以这种方式连接而成的链式分子就是核酸。无论是dna还是rna,其基本结构都是如此,故又称dna链或rna链。dna链的结构如下示意图。
寡核苷酸(oligonucleotide):这是与核酸有关的文献中经常出现的一个术语,一般是指二至十个核苷酸残基以磷酸二酯键连接而成的线性多核苷酸片段。但在使用这一术语时,对核苷酸残基的数目并无严格规定,在不少文献中,把含有三十甚至更多个核苷酸残基的多核苷酸分子也称作寡核苷酸。寡核苷酸可由仪器自动合成,它可作为dna合成的引物(primer)、基因探针(probe)等,在现代分子生物学研究中具有广泛的用途。
核酸链的简写式:核酸分子的简写式是为了更简单明了的叙述高度复杂的核酸分子而使用的一些简单表示式。它所要表示的主要内容是核酸链中的核苷酸(或碱基)。下面介绍二种常用的简写式。
字符式:书写一条多核苷酸链时,用英文大写字母缩写符号代表碱基(dna和rna中所含主要碱基及缩写符号见表1-1),用小写英文字母p代表磷酸残基。核酸分子中的糖基、糖苷键和酯键等均省略不写,将碱基和磷酸相间排列即可。因省略了糖基,故不再注解“脱氧”与否,凡简写式中出现t就视为dna链,出现u则视为rna链。以5'和3'表示链的末端及方向,分别置于简写式的左右二端。下面是分别代表dna链和rna链片段的二个简写式:
5'papcptptpgpapapcpg3'dna
5'papcpupupgpapapcpg3'rna
此式可进一步简化为:
5'pacttgaacg3'
5'pacuugaacg3'
上述简写式的5'-末端均含有一个磷酸残基(与糖基的c-5'位上的羟基相连),3'-末端含有一个自由羟基(与糖基的c-3'位相连),若5'端不写p,则表示5'-末端为自由羟基。双链dna分子的简写式多采用省略了磷酸残基的写法,在上述简式的基础上再增加一条互补链(complentarystrand)即可,链间的配对碱基用短纵线相连或省略,错配(mismatch)碱基对错行书写在互补链的上下两边,如下所示:
5'ggaatctcat3'
3'ccttagagta5'
5'ggaatc错配)
线条式:在字符书写基础上,以垂线(位于碱基之下)和斜线(位于垂线与p之间)分别表示糖基和磷酸酯键。
上式中,斜线与垂线部的交点为糖基的c-3'位,斜线与垂线下端的交点为糖基的c-5'位。这一书写式也可用于表示短链片段。不难看出,简写式表示的中心含义就是核酸分子的一级结构,即核酸分子中的核苷酸(或碱基)排列顺序。
分解代谢 编辑本段
从前面的描述我们也可以看得很清楚,核酸氧化分解后变成了磷酸和碱基的嘌呤和嘧啶,嘌呤也是导致人类尿酸增高和痛风的主要原因。
核酸氧化分解—生成嘌呤—嘌呤在肝脏进一步氧化成为(2,6,8—三氧嘌呤)又称为尿酸,尿酸盐沉积到关节腔等组织引起痛风发作。
因此,核酸不是越多越好,同时,这也说明了为什么中老年易患痛风,因为年纪大了,大量的细胞死亡,而细胞内有大量的核酸,生成嘌呤,再生成尿酸,从而导致痛风发作。防治好痛风就是要防止核酸被氧化。
相关分类 编辑本段
核酸(nucleic acid)是重要的生物大分子,它的构件分子是核苷酸(nucleotide)。
天然存在的核酸可分为:
⑴脱氧核糖核酸(deoxyribonucleic acid,dna)和核糖核酸(ribonucleic acid,rna)
dna贮存细胞所有的遗传信息,是物种保持进化和世代繁衍的物质基础。
rna中参与蛋白质合成的有三类:
转移rna(transfer rna,trna)、核糖体rna(ribosomal rna,rrna)和信使rna(messenger rna,mrna)
20世纪末,发现许多新的具有特殊功能的rna,几乎涉及细胞功能的各个方面。
核苷酸可分为:核糖核苷酸(rna的构件分子)和 脱氧核糖核苷酸(dna构件分子)
细胞内还有各种游离的核苷酸和核苷酸衍生物,它们具有重要的生理功能。
核苷酸由 核苷(nucleoside)磷酸(phosphonic.acid)组成
核苷由:碱基(base)和 戊糖(pentose)组成
构成核苷酸中的碱基是含氮杂环化合物,由嘧啶(pyrimidine)和嘌呤(purine)构成。
核酸:1.嘌呤碱:腺嘌呤(a)鸟嘌呤(g)2.嘧啶碱:胞嘧啶(c)胸腺嘧啶(t) 尿嘧啶(u)
dna中含有4种碱基:腺嘌呤、鸟嘌呤和胞嘧啶,胸腺嘧啶主要存在于dna中。
rna中含也有4种碱基:腺嘌呤、鸟嘌呤和胞嘧啶,尿嘧啶主要存在于rna中。
在某些trna分子中也有胸腺嘧啶,少数几种噬菌体的dna含尿嘧啶而不是胸腺嘧啶。这五种碱基受介质ph的影响出现酮式、烯醇式互变异构体。
在dna和rna中,尤其是trna中还有一些含量甚少的碱基,称为稀有碱基(rare bases)稀有碱基种类很多,大多数是甲基化碱基。trna中含稀有碱基高达10%。
核酸中有两种戊糖dna中为d-2-脱氧核糖(d-2-deoxyribose),rna中则为d-核糖(d-ribose)。在核苷酸中,为了与碱基中的碳原子编号相区别核糖或脱氧核糖中碳原子标以c-1’,c-2’等。脱氧核糖与核糖两者的差别只在于脱氧核糖中与2’位碳原子连结的不是羟基而是氢,这一差别使dna在化学上比rna稳定得多。
核苷是戊糖与碱基之间以糖苷键(glycosidic bond)相连接而成。戊糖中c-1’与嘧啶碱的n-1或者与嘌吟碱的n9相连接,戊糖与碱基间的连接键是n-c键,一般称为n-糖苷键。
rna中含有稀有碱基,并且还存在异构化的核苷。如在trna和rrna中含有少量假尿嘧啶核苷(用ψ表示),在它的结构中戊糖的c-1不是与尿嘧啶的n-1相连接,而是与尿嘧啶c-5相连接。
核苷中的戊糖5’碳原子上羟基被磷酸酯化形成核苷酸。核苷酸分为核糖核苷酸与脱氧核糖核苷酸两大类。依磷酸基团的多少,有一磷酸核苷、二磷酸核苷、三磷酸核苷。核苷酸在体内除构成核酸外,尚有一些游离核苷酸参与物质代谢、能量代谢与代谢调节,如三磷酸腺苷(atp)是体内重要能量载体;三磷酸尿苷参与糖原的合成;三磷酸胞苷参与磷脂的合成;环腺苷酸(camp)和环鸟苷酸(cgmp)作为第二信使,在信号传递过程中起重要作用;核苷酸还参与某些生物活性物质的组成:如尼克酰胺腺嘌呤二核苷酸(nad ),尼克酰胺腺嘌呤二核苷酸磷酸(nadp )和黄素腺嘌呤二核苷酸(fad)。
分子结构 编辑本段
核酸是由核苷酸聚合而成的生物大分子。组成dna的脱氧核糖核苷酸主要是damp、dgmp、dcmp和dtmp,组成rna的核糖核苷酸主要是amp、gmp、cmp和ump。核酸中的核苷酸以3’,5’磷酸二酯键构成无分支结构的线性分子。核酸链具有方向性,有两个末端分别是5’末端与3’末端。5’末端含磷酸基团,3’末端含羟基。核酸链内的前一个核苷酸的3’羟基和下一个核苷酸的5’磷酸形成3’,5’磷酸二酯键,故核酸中的核苷酸被称为核苷酸残基。通常将小于50个核苷酸残基组成的核酸称为寡核苷酸(oligonucleotide),大于50个核苷酸残基称为多核苷酸(polynucleotide)。
(一)dna的二级结构
dna二级结构即双螺旋结构(double helix structure)。20世纪50年代初chargaff等人分析多种生物dna的碱基组成发现的规则。
dna双螺旋模型的提出不仅揭示了遗传信息稳定传递中dna半保留复制的机制,而且是分子生物学发展的里程碑。
dna双螺旋结构特点如下:①两条dna互补链反向平行。②由脱氧核糖和磷酸间隔相连而成的亲水骨架在螺旋分子的外侧,而疏水的碱基对则在螺旋分子内部,碱基平面与螺旋轴垂直,螺旋旋转一周正好为10个碱基对,螺距为3.4nm,这样相邻碱基平面间隔为0.34nm并有一个36°的夹角。③dna双螺旋的表面存在一个大沟(major groove)和一个小沟(minor groove),蛋白质分子通过这两个沟与碱基相识别。④两条dna链依靠彼此碱基之间形成的氢键而结合在一起。根据碱基结构特征,只能形成嘌呤与嘧啶配对,即a与t相配对,形成2个氢键;g与c相配对,形成3个氢键。因此g与c之间的连接较为稳定。⑤dna双螺旋结构比较稳定。维持这种稳定性主要靠碱基对之间的氢键以及碱基的堆集力(stacking force)。
生理条件下,dna双螺旋大多以b型形式存在。右手双螺旋dna除b型外还有a型、c型、d型、e型。此外还发现左手双螺旋z型dna。z型dna是1979年rich等在研究人工合成的cgcgcg的晶体结构时发现的。z-dna的特点是两条反向平行的多核苷酸互补链组成的螺旋呈锯齿形,其表面只有一条深沟,每旋转一周是12个碱基对。研究表明在生物体内的dna分子中确实存在z-dna区域,其功能可能与基因表达的调控有关。dna二级结构还存在三股螺旋dna,三股螺旋dna中通常是一条同型寡核苷酸与寡嘧啶核苷酸-寡嘌呤核苷酸双螺旋的大沟结合,三股螺旋中的第三股可以来自分子间,也可以来自分子内。三股螺旋dna存在于基因调控区和其他重要区域,因此具有重要生理意义。
(二)dna三级结构——超螺旋结构
dna三级结构是指dna链进一步扭曲盘旋形成超螺旋结构。生物体内有些dna是以双链环状dna形式存在,如有些病毒dna,某些噬菌体dna,细菌染色体与细菌中质粒dna,真核细胞中的线粒体dna、叶绿体dna都是环状的。环状dna分子可以是共价闭合环,即环上没有缺口,也可以是缺口环,环上有一个或多个缺口。在dna双螺旋结构基础上,共价闭合环dna(covalently close circular dna)可以进一步扭曲形成超螺旋形(super helical form)。根据螺旋的方向可分为正超螺旋和负超螺旋。正超螺旋使双螺旋结构更紧密,双螺旋圈数增加,而负超螺旋可以减少双螺旋的圈数。几乎所有天然dna中都存在负超螺旋结构。
(三)dna的四级结构——dna与蛋白质形成复合物
在真核生物中其基因组dna要比原核生物大得多,如原核生物大肠杆菌的dna约为4.7×10kb,而人的基因组dna约为3×10 kb,因此真核生物基因组dna通常与蛋白质结合,经过多层次反复折叠,压缩近10 000倍后,以染色体形式存在于平均直径为5μm的细胞核中。线性双螺旋dna折叠的第一层次是形成核小体(nucleosome)。犹如一串念珠,核小体由直径为11nm×5.5nm的组蛋白核心和盘绕在核心上的dna构成。核心由组蛋白h2a、h2b、h3和h4各2分子组成,为八聚体,146 bp长的dna以左手螺旋盘绕在组蛋白的核心1.75圈,形成核小体的核心颗粒,各核心颗粒间有一个连接区,约有60 bp双螺旋dna和1个分子组蛋白h1构成。平均每个核小体重复单位约占dna 200 bp。dna组装成核小体其长度约缩短7倍。在此基础上核小体又进一步盘绕折叠,最后形成染色体。
(四)dna结构的多态性
watson和crick所推导出来的dna结构在生物学研究中有深远意义。他们是以在生理盐溶液中抽出的dna纤维在92%相对温度下进行x-射线衍射图谱为依据进行推设的。在这一条件下得出的dna称b构象。实际上在溶液中的dna的确呈这一构象,这也是最常见的dna构象。但是,研究表明dna的结构是动态的。在以钠、钾或铯作反离子,相对温度为75%时,dna分子的x-射线衍射图给出的是a构象。这一构象不仅出现于脱水dna中,还出现在rna分子中的双螺旋区域的dna-rna杂交分子中。如果以锂作反离子,相对温度进一步降为66%,则dna是c构象。但是这一构象仅在实验室中观察到,还未在生物体中发现。这些dna分子中g-c碱基对较少,这些分子将取d和e构象。这些研究表明dna的分子结构不是一成不变的,在不同的条件下可以有所不同。但是,这些不同构象的dna都有共同的一点,即它们都是右手双螺旋;两条反向平行的核苷酸链通过watson-crick碱基配对结合在一起;链的重复单位是单核苷酸;这些螺旋中都有两个螺旋沟,分为大沟与小沟,只是它们的宽窄和深浅程度有所不同。
但是,wang和rich等人在研究人工合成的cgcgcg单晶的x-射线衍射图谱时分别发现这种六聚体的构象与上面讲到的完全不同。它是左手双螺旋,在主链中各个磷酸根呈锯齿状排列,有如“之”字形一样,因此叫它z构象(英文字zigzag的第一个字母)。还有,这一构象中的重复单位是二核苷酸而不是单核苷酸;而且z-dna只有一个螺旋沟,它相当于b构象中的小沟,它狭而深,大沟则不复存在。
立即就有几个问题被提了出来:这种结构是怎样生成的?这一结构在天然状态下存在吗?它有什么生物学意义?
研究表明,z-dna的形成是dna单链上出现嘌呤与嘧啶交替排列所成的。比如cgcgcgcg或者cacacaca。这种碱基排列方式会造成核苷酸的糖苷键的顺式和反式构象的交替存在。当碱基与糖构成反式结构时,它们之间离得远;而当它们成顺式时,就彼此接近。嘧啶糖苷键通常是反式的,而嘌呤糖苷酸键既可成顺式的也可成反式的。而在z-dna中,嘌呤碱是顺式的。这样,在z-dna中嘧啶的糖苷链离开小沟向外挑出,而嘌呤上的糖苷键则弯向小沟。嘌呤与嘧啶的交替排列就使得糖苷键也是顺式与反式交替排列,从而使z-dna主链呈锯齿状或“之”字形。
人们相信,并用实验证明细胞dna分子中确实存在有z-dna区。而且,细胞内有一些因素可以促使b-dna转变为z-dna。比如,胞嘧啶第五位碳原子的甲基化,在甲基周围形成局部的疏水区。这一区域扩伸到b-dna的大沟中,使b-dna不稳定而转变为z-dna。这种c5甲基化现象在真核生物中是常见的。因此在生物b构象的dna中某些区段具有z-dna构象是可能的。dna真是一个构象可变动态分子。
z-dna有会么生物学意义呢?应当指出z-dna的形成通常在热力学上是不利的。因为z-dna中带负电荷的磷酸根距离太近了,这会产物静电排斥。但是,dna链的局部不稳定区的存在就成为潜在的解链位点。dna解螺旋却是dna复制和转录等过程中必要的环节,因此认为这一结构位点与基因调节有关。比如sv40增强子区中就有这种结构,又如鼠类微小病毒dns复制区起始点附近有gc交替排列序列。此外,dna螺旋上沟的特征在其信息表达过程中起关键作用。调控蛋白都是通过其分子上特定的氨基酸侧链与dna双螺旋沟中的碱基对一侧的氢原子供体或受体相互作用,形成氢键从而识别dna上的遗传信息的。大沟所带的遗传信息比小沟多。沟的宽窄和深浅也直接影响到调控蛋白质对dna信息的识别。z-dna中大沟消失,小沟狭而深,使调探蛋白识别方式也发生变化。这些都暗示z-dna的存在不仅仅是由于dna中出现嘌呤-啶嘧交替排列之结果,也一定是在漫漫的进化长河中对dna序列与结构不断调整与筛选的结果,有其内在而深刻的含意,只是人们还未充分认识而已。
dna构象的可变性,或者说dna二级结构的多态性的发现拓宽了人们的视野。原来,生物体中最为稳定的遗传物质也可以采用不同的姿态来实现其丰富多彩的生物的奥妙,也让人们在这一领域中探索和攀越时减少疲劳和厌倦,乐而忘返,从而有更多更新的发现。
多年来,dna结构的研究手段主要是x射线衍线技术,其结果是通过间接观测多个dna分子有关结构参数的平均值而获得的。同时,这项技术的样品分析条件使被测dna分子与天然状态相差甚远。因此,在反映dna结构真实性方面这种方法存在着缺陷。1989年,应用扫描隧道显微镜(stm)研究dna结构克服了上述技术的缺陷。这种先进的显微技术,不仅可将被测物放大500万倍,且能直接观测接近天然条件下单个dna分子的结构细节。应该说它所取得的dna结构资料更具有"权威性"。表1-6是stm测到的b-dna结构参数及其与x射线衍线资料的比较结果。stm研究还证实了d(cg)重复序列的寡核苷酸片段为z-dna结构的事实。stm技术的应用是dna结构研究中的重要进展,可望在探索dna结构的某些未知点上展示巨大潜力。
(一)基因(gene)的现代分子生物学概念是指能编码有功能的蛋白质多肽链或合成rna所必需的全部核酸序列,是核酸分子的功能单位。一个基因通常包括编码蛋白质多肽链或rna的编码序列,保证转录和加工所必需的调控序列和5’端、3’端非编码序列。另外在真核生物基因中还有内含子等核酸序列。
(二)基因组(genome)是指一个细胞或病毒所有基因及间隔序列,储存了一个物种所有的遗传信息。在病毒中通常是一个核酸分子的碱基序列,单细胞原核生物是它仅有的一条染色体的碱基序列,而多细胞真核生物是一个单倍体细胞内所有的染色体。如人单倍体细胞的23条染色体的碱基序列。多细胞真核生物起源于同一个受精卵,其每个体细胞的基因组都是相同的。
病毒基因组 famous encyclopedia string
原核生物基因组
famous encyclopedia string
真核生物基因组 famous encyclopedia string
在高等真核生物中基因序列占整个基因组不到10%,大部分是非编码的间隔序列。人类基因组研究结果发现在人的基因组中与蛋白质合成有关的基因只占整个基因组2 %。真核生物基因组的最大的特点是出现分隔开的基因,在这类基因中有编码作用的序列称外显子(exon),没有编码作用的序列称内含子(intron),它们彼此间隔排列。
绝大部分rna分子都是线状单链,但是rna分子的某些区域可自身回折进行碱基互补配对,形成局部双螺旋。在rna局部双螺旋中a与u配对、g与c配对,除此以外,还存在非标准配对,如g与u配对。rna分子中的双螺旋与a型dna双螺旋相似,而非互补区则膨胀形成凸出(bulge)或者环(loop),这种短的双螺旋区域和环称为发夹结构(hairpin)。发夹结构是rna中最普通的二级结构形式,二级结构进一步折叠形成三级结构,rna只有在具有三级结构时才能成为有活性的分子。rna也能与蛋白质形成核蛋白复合物,rna的四级结构是rna与蛋白质的相互作用。
(一) trna的结构
trna约占总rna的15%,trna主要的生理功能是在蛋白质生物合成中转运氨基酸和识别密码子,细胞内每种氨基酸都有其相应的一种或几种trna,因此trna的种类很多,在细菌中约有30~40种trna,在动物和植物中约有50~100种trna。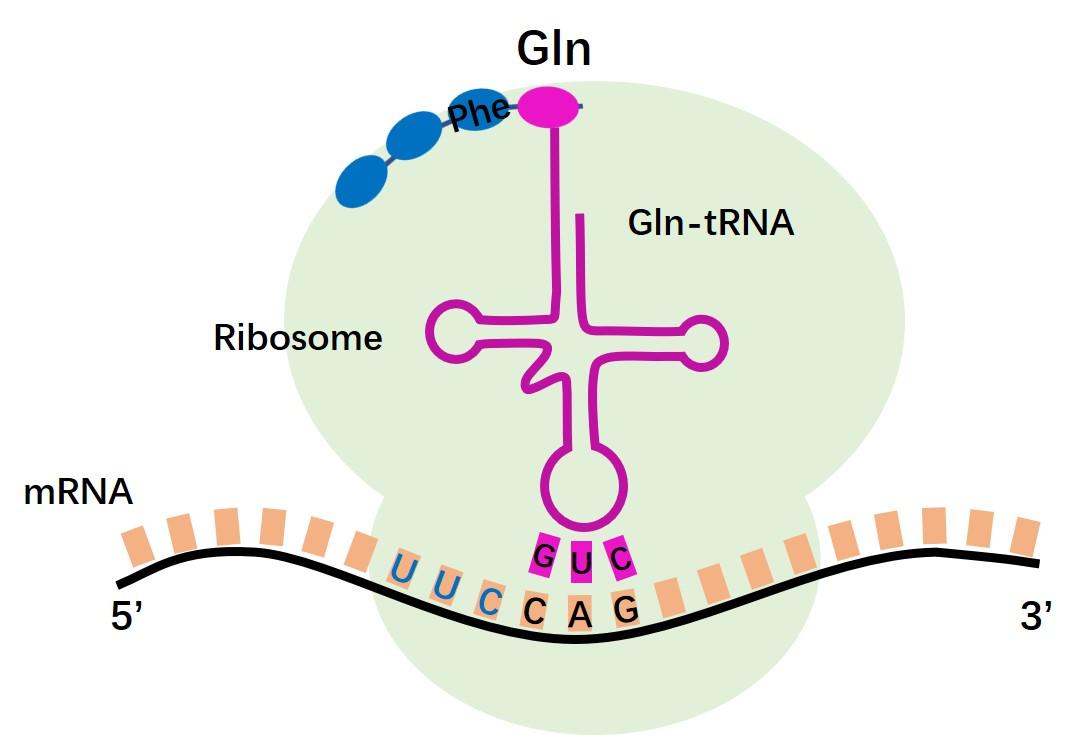 1. trna一级结构:
1. trna一级结构:
trna是单链分子,含73~93核苷酸,分子质量为24 000~31 000kda,沉降系数4s。含有10%的稀有碱基。如二氢尿嘧啶(dhu)、核糖胸腺嘧啶(rt)和假尿苷(ψ)以及不少碱基被甲基化,其3’端为cca-oh,5’端多为pg,分子中大约30%的碱基是不变的或半不变的,也就是说它们的碱基类型是保守的。
2. trna二级结构: trna二级结构为三叶草型(如右图)。配对碱基形成局部双螺旋而构成臂,不配对的单链部分则形成环。三叶草型结构由4臂4环组成。氨基酸臂由7对碱基组成,双螺旋区的3’末端为一个4个碱基的单链区-ncca-oh 3’,腺苷酸残基的羟基可与氨基酸α羧基结合而携带氨基酸。二氢尿嘧啶环以含有2个稀有碱基二氢尿嘧啶(dhu)而得名,不同trna其大小并不恒定,在8~14个碱基之间变动,二氢尿嘧啶臂一般由3~4对碱基组成。反密码环由7个碱基组成,大小相对恒定,其中3个核苷酸组成反密码子(anticodon),在蛋白质生物合成时,可与mrna上相应的密码子配对。反密码臂由5对碱基组成。额外环在不同trna分子中变化较大可在4~21个碱基之间变动,又称为可变环,其大小往往是trna分类的重要指标。tψc环含有7个碱基,大小相对恒定,几乎所有的trna在此环中都含tψc序列,tψc臂由5对碱基组成。
3. trna的三级结构:
二十世纪七十年代初科学家用x线射衍技术分析发现trna的三级结构为倒l形(如右图)。trna三级结构的特点是氨基酸臂与tψc臂构成l的一横,-ccaoh3’末端就在这一横的端点上,是结合氨基酸的部位,而二氢尿嘧啶臂与反密码臂及反密码环共同构成l的一竖,反密码环在一竖的端点上,能与mrna上对应的密码子识别,二氢尿嘧啶环与tψc环在l的拐角上。形成三级结构的很多氢键与trna中不变的核苷酸密切有关,这就使得各种trna三级结构都呈倒l形的。在trna中碱基堆积力是稳定trna构型的主要因素。
(二)mrna
原核生物中mrna转录后一般不需加工,直接进行蛋白质翻译。mrna转录和翻译不仅发生在同一细胞空间,而且这两个过程几乎是同时进行的。真核细胞成熟mrna是由其前体核内不均一rna(heterogeneous nuclear rna,hnrna)剪接并经修饰后才能进入细胞质中参与蛋白质合成。所以真核细胞mrna的合成和表达发生在不同的空间和时间。mrna的结构在原核生物中和真核生物中差别很大。下面分别作一介绍:
1. 原核生物mrna结构特点
原核生物的mrna结构简单,往往含有几个功能上相关的蛋白质的编码序列,可翻译出几种蛋白质,为多顺反子。在原核生物mrna中编码序列之间有间隔序列,可能与核糖体的识别和结合有关。在5’端与3’端有与翻译起始和终止有关的非编码序列,原核生物mrna中没有修饰碱基,5’端没有帽子结构,3’端没有多聚腺苷酸的尾巴(polyadenylate tail,polya尾巴)。原核生物的mrna的半衰期比真核生物的要短得多,转录后1min,mrna降解就开始。
2. 真核生物mrna结构特点
真核生物mrna为单顺反子结构,即一个mrna分子只包含一条多肽链的信息。在真核生物成熟的mrna中5’端有m7gpppn的帽子结构,帽子结构可保护mrna不被核酸外切酶水解,并且能与帽结合蛋白结合识别核糖体并与之结合,与翻译起始有关。3’端有polya尾巴,其长度为20~250个腺苷酸,其功能可能与mrna的稳定性有关,少数成熟mrna没有polya尾巴,如组蛋白mrna,它们的半衰期通常较短。
(三)rrna的结构
rrna占细胞总rna的80%左右,rrna分子为单链,局部有双螺旋区域具有复杂的空间结构,原核生物主要的rrna有三种,即5s、16s和23s rrna,如大肠杆菌的这三种rrna分别由120、1542和2904个核苷酸组成。真核生物则有4种,即5s、5.8s、18s和28s rrna,如小鼠,它们相应含121、158、1874和4718个核苷酸。rrna分子作为骨架与多种核糖体蛋白(ribosomal protein)装配成核糖体。
所有生物体的核糖体都由大小不同的两个亚基所组成。原核生物核糖体为70s,由50s和30s两个大小亚基组成。30s小亚基含16s的rrna和21种蛋白质,50s大亚基含23s和5s两种rrna及34种蛋白质。真核生物核糖体为80s,是由60s和40s两个大小亚基组成。40s的小亚基含18s rrna及33种蛋白质,60s大亚基则由28s、5.8s和5s 3种rrna及49种蛋白质组成。
(四)其他rna分子
20世纪80年代以后由于新技术不断产生,人们发现rna有许多新的功能和新的rna基因。细胞核内小分子rna(small nuclear rna,snrna)是细胞核内核蛋白颗粒(small nuclear ribonucleoprotein particles,snrnps)的组成成分,参与mrna前体的剪接以及成熟的mrna由核内向胞浆中转运的过程。核仁小分子rna(small nucleolar rna,snorna)是类新的核酸调控分子,参与rrna前体的加工以及核糖体亚基的装配。胞质小分子rna(small cytosol rna, scrna)的种类很多,其中7s lrna与蛋白质一起组成信号识别颗粒(signal recognition particle,srp),srp参与分泌性蛋白质的合成,反义rna(antisense rna)由于它们可以与特异的mrna序列互补配对,阻断mrna翻译,能调节基因表达。核酶是具有催化活性的rna分子或rna片段。在医学研究中已设计了针对病毒的致病基因mrna的核酶,抑制其蛋白质的生物合成,为基因治疗开辟新的途径,核酶的发现也推动了生物起源的研究。微rna(microrna,mirna)是一种具有茎环结构的非编码rna,长度一般为20-24个核苷酸,在mrna翻译过程中起到开关作用,它可以与靶mrna结合,产生转录后基因沉默作用(post-transcriptional gene silencing,ptgs),在一定条件下能释放,这样mrna又能翻译蛋白质,由于mirna的表达具有阶段特异性和组织特异性,它们在基因表达调控和控制个体发育中起重要作用。
随着基因组研究不断深入,蛋白组学研究逐渐展开,rna的研究也取得了突破性的进展,发现了许多新的rna分子,人们逐渐认识到dna是携带遗传信息分子,蛋白质是执行生物学功能分子,而rna即是信息分子,又是功能分子。人类基因组研究结果表明,在人类基因组中约有30000~40000个基因,其中与蛋白质生物合成有关的基因只占整个基因组的2%,对不编码蛋白质的98%基因组的功能有待进一步研究,为此20世纪末科学家在提出蛋白质组学后,又提出rna组学。rna组是研究细胞的全部rna基因和rna的分子结构与功能。
相关性质 编辑本段
①酸效应:在强酸和高温,核酸完全水解为碱基,核糖或脱氧核糖和磷酸。在浓度略稀的的无机酸中,最易水解的化学键被选择性的断裂,一般为连接嘌呤和核糖的糖苷键,从而产生脱嘌呤核酸。
②碱效应
1. dna:当ph值超出生理范围(ph7~8)时,对dna结构将产生更为微妙的影响。碱效应使碱基的互变异构态发生变化。这种变化影响到特定碱基间的氢键作用,结果导致dna双链的解离,称为dna的变性
2.rna:ph较高时,同样的变性发生在rna的螺旋区域中,但通常被rna的碱性水解所掩盖。这是因为rna存在的2`-oh参与到对磷酸脂键中磷酸分子的分子内攻击,从而导致rna的断裂。
③化学变性:一些化学物质能够使dna/rna在中性ph下变性。由堆积的疏水碱基形成的核酸二级结构在能量上的稳定性被削弱,则核酸变性。
①黏性:dna的高轴比等性质使得其水溶液具有高黏性,很长的dna分子又易于被机械力或超声波损伤,同时黏度下降。
② 浮力密度:可根据dna的密度对其进行纯化和分析。在高浓度分子质量的盐溶液(cscl)中,dna具有与溶液大致相同的密度,将溶液高速离心,则cscl趋于沉降于底部,从而建立密度梯度,而dna最终沉降于其浮力密度相应的位置,形成狭带,这种技术成为平衡密度梯度离心或等密度梯度离心。
③稳定性:核酸的结构相当稳定,其主要原因有1、碱基对间的氢键2、碱基的堆积作用3、环境中的阳离子。
①减色性:dsdna相对于ssdna是减色的,而ssdna相对于dsdna是增色的。
② dna纯度:a260/a280。
①热变性:dsdna与rna的热力学表现不同,随着温度的升高rna中双链部分的碱基堆积会逐渐地减少,其吸光性值也逐渐地,不规则地增大。较短的碱基配对区域具有更高的热力学活性,因而与较长的区域相比变性快。而dsdna热变性是一个协同过程。分子末端以及内部更为活跃的富含a-t的区域的变性将会使其赴京的螺旋变得不稳定,从而导致整个分子结构在解链温度下共同变性。
② 复性:dna的热变性可通过冷却溶液的方法复原。不同核酸链之间的互补部分的复性称为杂交。
一般来说,进化程度高的生物dna分子应越大,能贮存更多遗传信息。但进化的复杂程度与dna大小并不完全一致,如哺乳类动物dna约为3×10 bp,但有些两栖类动物、南美肺鱼dna大小可达1010bp到1011bp。
常用测定dna分子大小的方法有电泳法、离心法。凝胶电泳是当前研究核酸的最常用方法,凝胶电泳有琼脂糖(agarose)凝胶电泳和聚丙烯酰胺(polyacrylamide)凝胶电泳。
dna和rna中的糖苷键与磷酸酯键都能用化学法和酶法水解。在很低ph条件下dna和rna都会发生磷酸二酯键水解。并且碱基和核糖之间的糖苷键更易被水解,其中嘌呤碱的糖苷键比嘧啶碱的糖苷键对酸更不稳定。在高ph时,rna的磷酸酯键易被水解,而dna的磷酸酯键不易被水解。
水解核酸的酶有很多种,若按底物专一性分类,作用于rna的称为核糖核酸酶(ribonuclease,rnase),作用于dna的则称为脱氧核糖核酸酶(deoxyribonuclease,dnase)。按对底物作用方式分类,可分核酸内切酶(endonuclease)与核酸外切酶(exonuclease)。核酸内切酶的作用是在多核苷酸内部的3’,5’磷酸二酯键,有些内切酶能识别dna双链上特异序列并水解有关的3’,5’磷酸二酯键。核酸内切酶是非常重要的工具酶,在基因工程中有广泛用途。而核酸外切酶只对核酸末端的3’,5’磷酸二酯键有作用,将核苷酸一个一个切下,可分为5’→3’外切酶,与3’→5’外切酶。
在一定理化因素作用下,核酸双螺旋等空间结构中碱基之间的氢键断裂,变成单链的现象称为变性(denaturation)。引起核酸变性的常见理化因素有加热、酸、碱、尿素和甲酰胺等。在变性过程中,核酸的空间构象被破坏,理化性质发生改变。由于双螺旋分子内部的碱基暴露,其a260值会大大增加。a260值的增加与解链程度有一定比例关系,这种关系称为增色效应(hyperchromic effect)。如果缓慢加热dna溶液,并在不同温度测定其a260值,可得到“s”形dna熔化曲线(melting curve)。从dna熔化曲线可见dna变性作用是在一个相当窄的温度内完成的。
当a260值开始上升前dna是双螺旋结构,在上升区域分子中的部分碱基对开始断裂,其数值随温度的升高而增加,在上部平坦的初始部分尚有少量碱基对使两条链还结合在一起,这种状态一直维持到临界温度,此时dna分子最后一个碱基对断开,两条互补链彻底分离。通常把加热变性时dna溶液a260升高达到最大值一半时的温度称为该dna的熔解温度(melting temperature tm),tm是研究核酸变性很有用的参数。tm一般在85~95℃之间,tm值与dna分子中g c含量成正比。
变性dna在适当条件下,可使两条分开的单链重新形成双螺旋dna的过程称为复性(renaturation)。当热变性的dna经缓慢冷却后复性称为退火(annealing)。dna复性是非常复杂的过程,影响dna复性速度的因素很多:dna浓度高,复性快;dna分子大复性慢;高温会使dna变性,而温度过低可使误配对不能分离等等。最佳的复性温度为tm减去25℃,一般在60℃左右。离子强度一般在0.4mol/l以上。
具有互补序列的不同来源的单链核酸分子,按碱基配对原则结合在一起称为杂交(hybridization)。杂交可发生在dna-dna、rna-rna和dna-rna之间。杂交是分子生物学研究中常用的技术之一,利用它可以分析基因组织的结构,定位和基因表达等,常用的杂交方法有southern印迹法,northern印迹法和原位杂交(insitu hybridization)等。
区别对比 编辑本段
核酸、蛋白质谁更“牛”?
一般人都知道,生命是蛋白质存在的形式,蛋白质是生命的基础。在发现核酸前,这句话是对的,但当核酸被发现后,应该说最本质的生命物质是核酸,或是把上述的这句话更正为核酸是生命的基础。按照现代生物学的观点,蛋白体是包括核酸和蛋白质的生物大分子。
核酸在生命中为什么比蛋白质更重要呢?因为生命的重要性是能自我复制,而核酸就能够自我复制。蛋白质的复制是根据核酸所发出的指令,使氨基酸根据其指定的种类进行合成,然后再按指定的顺序排列成所需要复制的蛋白质。世界上各种有生命的物质都含有蛋白体,蛋 白体中有核酸和蛋白质,还没有发现有蛋白质而没有核酸的生命。但在有生命的病毒研究中,却发现病毒以核酸为主体,蛋白质和脂肪以及脂蛋白等只不过充作其外壳,作为与外界环境的界限而已,当它钻入寄生细胞繁殖子代时,把外壳留在细胞外,只有核酸进入细胞内,并使细胞在核酸控制下为其合成子代的病毒。这种现象,美国科学家比喻为人和汽车的关系。即把核酸比为人,蛋白质比作汽车,人驾驶汽车到处跑,外表上看,人车一体是有生命运动的东西,而真正的生命是人,汽车只是由人制造的载入的外壳。有一种类病毒,是能繁殖子代的有生命物体,其中只有核酸而没蛋白质,可见核酸是真正的生命物质。
因此中国1996年最新出版的《人体生理学》改变了旧教科书中只提蛋白质是生命基础的缺陷,明确提出:“蛋白质和核酸是一切生命活动的物质基础。”
然而,多少年来,人们在一味追求蛋白质、维生素、微量元素等营养时,却把最重要的角色——核酸忘却了,这不能不说是人类生命史上的一大遗憾。
没有核酸,就没有蛋白,也就没有生命。
核酸也称多聚核苷酸,是由许多个核苷酸聚合而成的生物大分子,核苷酸是由含氮的碱基、核糖或脱氧核糖、磷酸三种分子连接而成。碱基与糖通过糖苷键连接成核苷,核苷与磷酸以酯键连接成核苷酸。核苷酸是生物体内一类重要含氮化合物,是各种核酸的基本组成单位。根据核酸所含戊糖的不同,可分为核糖核酸(rna)和脱氧核糖核酸(dna)二种。
核酸不但是一切生物细胞的基本成分,还对生物体的生长、发育、繁殖、遗传及变异等重大生命现象起主宰作用。它在生物科学的地位,可用“没有核酸就没有生命”这句话来概括。
饮食核酸的营养保健作用如下:
1.饮食核酸与免疫 ‘
从核酸对机体各系统的影响来看,免疫系统是最敏感也是最直接受影响的系统。1985年科学家就证实无核酸饮食或低核酸饮食配方饲喂的实验动物,其细胞免疫功能低下,条件致病菌就可使其感染。无核酸饮食致使t淋巴细胞发育障碍、功能低下,而没有细胞免疫反应的发生,同时影响t细胞依赖的体液免疫的产生;补充核酸营养后可恢复免疫系统的发育和免疫功能。实验表明,核酸是维持机体正常免疫功能和免疫系统生长代谢的必需营养物质。
2.饮食核酸与衰老和内分泌
衰老是机体各组织器官的退行性变化,关于衰老发生机制的学说很多,如自由基学说、免疫学说、内分泌学说、遗传学说等。脂质过氧化随年龄增大而增高,并伴有酶与非酶系统防御功能下降,导致体内自由基浓度升高。代谢性、退行性疾病的发生和发展与体内过氧化脂质含量高度正相关。饮食核酸作为使遗传物质活泼代谢的原料,具有极强的抗生物氧化、消除体内自由基和全面增强免疫功能及性激素分泌的作用,因此在延缓衰老方面优势显著。
3.饮食核酸与增殖细胞
饮食中补加核酸有助于肝脏再生和受损伤的小肠恢复功能。有无核酸饮食对比研究证明,一段时期内膳食中如缺乏核酸,将对大鼠肝脏的超微结构及功能造成不良影响,提示饮食核酸是维持肝脏处于正常生理状态的必需营养物质。血液中的红细胞、白细胞、血小板和血浆蛋白等也都是代谢较快的增殖细胞系,加之它们中的大多数均无从头合成核酸的能力,因此它们的代谢和功能也都需要充足的核酸营养。再生障碍性贫血和抗癌药物、放疗、化疗等引起的贫血,即缺铁性贫血之外的贫血均需补充核酸营养,以改善骨髓造血功能和血液成分的代谢活力。
4.饮食核酸与癌症
人体每日约有数百万个癌状细胞出现,它们几乎全部被机体的免疫监视系统和核酸、维生素等食物成分,在形成大的癌细胞克隆前排除掉。因此在日常生活中尽量避免致癌因子的作用,增加核酸等防癌因素的作用非常必要。
5.饮食核酸与痴呆等神经障碍
食物核酸提取物对痴呆症状的改善非常令人鼓舞。在大鼠实验中,如给大鼠脑注射rna合成阻断剂,则所掌握的学习能力和记忆能力在5小时后丧失,但如在注射rna合成阻断剂的同时注射拮抗阻断剂的物质,这种记忆丧失就不发生。美国哈佛大学的研究也表明,老年痴呆患者脑内神经纤维变化多的部位,rna和蛋白质合成显著减少,因此发生记忆障碍。
6.饮食核酸与循环系统
核酸营养对循环系统的作用是抑制过氧化脂质的形成,抑制胆固醇的生成,扩张血管,改善血流,纠正心肌代偿不良,促进血管壁再生,抑制血小板凝集i因此核酸被认为对脑血栓、心肌梗死、高血压和动脉粥样硬化症有较好的营养保健作用。
7、饮食核酸与糖尿病
非胰岛素依赖性糖尿病与生活方式和运动不足关系密切,如果在普通的饮食疗法的基础上,再加上核酸饮食,将收到更好的效果。其原因:一是糖尿病患者血清中过氧化脂质增多,核酸及其代谢产物对其具有较强的清除作用;二是由于核酸的促细胞(包括促胰脏的胰岛素分泌细胞)代谢功能。除此之外,核酸的代谢产物腺苷还有抑制糖的分解作用,使糖在小肠内的吸收减缓。
除上述作用外,饮食核酸还有以下作用:减肥,提高机体对环境变化的耐受力,显著的抗疲劳、增强机体对冷热的抵抗力、促进摄人氧气的利用,促进小鼠生殖系统的发育等。
对于婴儿、迅速成长期的孩子、老年体弱多病、全身感染、外伤手术者、肝功能不全以及白细胞、t细胞、淋巴细胞降低人群等,可以额外补充核酸类物质。世界卫生组织规定,每天膳食中核酸的量不大于2克,扣除食物中的核酸摄入量,每天补充小于1.5克核酸是合适的。
人工核酸治疗白血病
日本工业技术院产业技术融合领域研究所已开发出了治疗白血病的人造核酸。科研小组的成员、东京大学研究生院教授多比良和诚根据动物实验结果认为,这种人造核酸将来有望成为治疗白血病的主要药物。
这次研究的对象是慢性骨髓性白血病(mcl),患者的异常遗传因子是由两个正常的遗传因子连接而成的,新开发的人造核酸可以发现这种变异遗传基因并将其切断。科学家过去也发现过能找到特定的遗传因子序列并将其切断的分子,但在切断特定遗传因子序列的同时往往对正常细胞造成伤害。而新开发出的核酸只在发现异常遗传因子时才被激活,平时则潜伏不动。
科研小组用人体白血病细胞进行了动物实验。他们将可与人造核酸反应的细胞和不可与人造核酸反应的细胞分别注射到8只实验鼠的体内。移植后第13周时,不与人造核酸反应的细胞全部死亡,而与人造核酸反应的细胞全部存活,证明人造核酸在生物体内十分有效。
科研小组说,此人造核酸的临床应用尚有诸多问题要解决,将来很可能是把患者的骨髓细胞抽出来,经人造核酸处理后,再把正常细胞的骨髓输回患者体内。
核酸的生物功能
核酸在生物体内主要与蛋白质合成核蛋白存在,它既是蛋白质生物合成不可缺少的物质,又是生物遗传的物质基础。
dna主要存在于细胞核中,它们是遗传信息的携带者,dna的结构决定生物合成蛋白质的特定结构,并保证这种遗传特性传给下一代。rna主要存在于细胞质中,它们是以dna为模板而形成的,并且直接参加蛋白质的生物合成过程。因此,dna是rna的模板,而rna又是蛋白质的模板。存在于dna分子上的遗传信息就是这样由dna传递给rna,再传递给蛋白质。通过dna 复制,遗传信息一代代传下去,正因为有这样的功能,人们把核酸誉为“生命之源”和“生命之本”。
附件列表
词条内容仅供参考,如果您需要解决具体问题
(尤其在法律、医学等领域),建议您咨询相关领域专业人士。